| Sistema Complemento | Lámina 3 |
|||
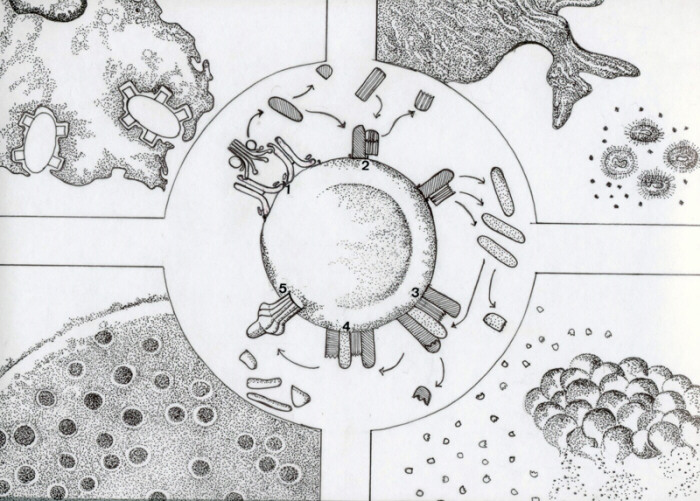
El Sistema Complemento está formado por un conjunto de proteínas séricas que se activan en cascada y por proteínas reguladoras. La activación puede iniciarse a través de complejos antígeno-anticuerpo en la vía clásica y por acción de componentes bacterianos y diversas enzimas en la vía alterna. Ambas vías convergen a nivel de C3 para originando el mismo producto final. La vía clásica (figura central) se inicia cuando C1q se une a fragmento Fc de dos moléculas de inmunoglobulinas unidas al antígeno presente en una membrana (1). Se produce la fragmentación de C1r y se activa C1s el cual fragmenta C4 y C2 originando C4b2b unido a membrana (2). C4a, que tiene actividad de anafilatoxina (induce liberación de mediadores en células cebadas y basófilos), y C2a difunden al fluído. El complejo bimolecular C4b2b es una C3 convertasa, esto es, rompe múltiples moléculas de C3 originando C3b que se une al complejo C4b2b para formar la C5 convertasa (3) y C3a que es una anafilatoxina. C3b cumple también la importante función de opsonina, ya que al unirse a la membrana, confiere mayor eficacia a la fagocitosis.
La C5 convertasa formada por C4b2b3b escinde la molécula C5 en los fragmentos C5b y C5a. C5a es anafilotoxina y C5b es una proteína de anclaje para C6. Posteriormente se unen a ellas C7 y C8 formando un complejo estable asociado a membrana que une a C9 (4). Este último se polimeriza formando junto con C5b, C6, C7 y C8 el complejo de ataque a membrana responsable de la lisis celular (5). En el sistema Complemento participan además de las proteínas nombradas, inhibidores específicos e inespecíficos y reguladores tanto solubles como constitutivos de membrana. Las principales funciones biológicas del Sistema Complemento incluyen la opsonización (A), la quimiotaxis (B), la lisis celular y bacteriana (C) y la función de anafilatoxina (D). Además participa en la eliminación de complejos inmunes. A. Opsonización: el factor C3b se une covalentemente a la superficie de bacterias o células constituyéndose en un ligando para un receptor específico ubicado en la membrana de polimorfonucleares y macrófagos entre otros. De esta manera, la bacteria queda anclada a la superficie celular facilitando la formación del fagosoma y estimulando el estallido metabólico que lleva a la producción de radicales libres derivados del oxígeno. Existen cuatro receptores para fragmentos de C3b en los leucocitos, CR1, CR2, CR3 y CR4, cuya deficiencia determina una mayor susceptibilidad a padecer infecciones recurrentes. B. Quimiotaxis: el fragmento C5a, en conjunto con leucotrienos y citoquinas ( IL-8 ) ejercen atracción sobre leucocitos y monocitos guiando su migración hacia el lugar donde se encuentra el agente injuriante. Además, al interactuar con receptores específicos en los fagocitos aumentan su motilidad y generan mecanismos bactericidas oxígeno-dependientes al activar su metabolismo oxidativo. C. Lisis: el complejo de ataque a membrana se introduce en la membrana de bacterias, células y algunos virus produciendo alteraciones de permeabilidad que conducen a su muerte. D. Anafilatoxina: C3a, C4a y C5a inducen la liberación de mediadores inflamatorios en diversas células los que producen el aumento de la permeabilidad vascular característico de la anafilaxia. Las células cebadas, basófilos, células musculares lisas y linfocitos expresan receptores para C3a y C4a. Las células cebadas, basófilos, neutrófilos, monocitos/macrófagos y endotelios poseen receptores para C5a. Las células cebadas y basófilos son estimuladas a liberar mediadores químicos especialemente histamina la que produce aumento de la permeabilidad vascular y contracción del músculo liso. Las células musculares y endoteliales se contraen por acción directa de las anafilatoxinas. |
||||