| Fagocitosis | Lámina 6 |
|||
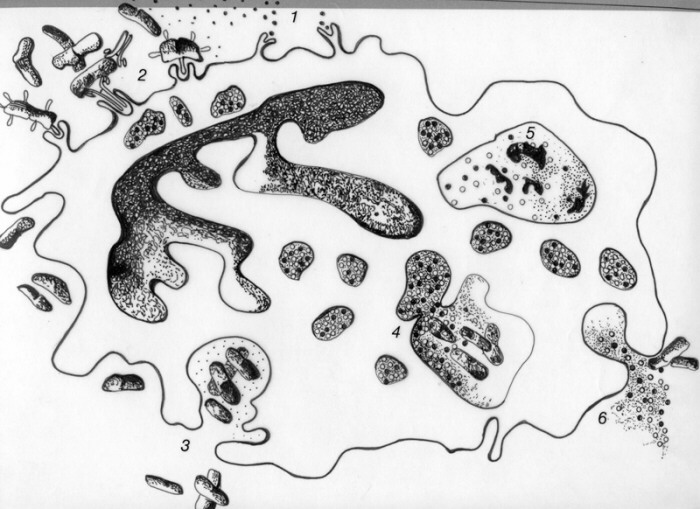
La fagocitosis es un mecanismo básico de defensa presente en la mayoría de las especies. En los mamíferos está a cargo de células especializadas, principalmente los polimorfonucleares neutrófilos y los macrófagos. Los primeros son células sanguíneas circulantes que se ponen en contacto con el material a fagocitar a través de la respuesta inflamatoria. Los macrófagos provienen de monocitos circulantes o bien están distribuidos estratégicamente en los tejidos. Ambos presentan diferencias en su poder fagocítico.
Los polimorfonucleares neutrófilos son especialmente efectivos durante el inicio de infecciones por bacterias extracelulares gram positivas y gram negativas. Los macrófagos participan en etapas más tardías de la inflamación fagocitando bacterias y restos celulares. Son importantes en la defensa ante bacterias intracelulares y generalmente actúan en concomitancia con respuestas inmunes adaptativas. La fagocitosis por polimorfonucleares neutrófilos se inicia a raiz del contacto de esta célula con factores quimiotácticos endógenos o elaborados por microorganismos (N-formilpéptidos)(1). Esta unión permite la migración direccional del fagocito hacia el agente injuriante y activa el metabolismo oxidativo celular. El reconocimiento y contacto con material a ser fagocitado está mediado por opsoninas que pueden ser anticuerpos o el fragmento C3b del complemento (2). La unión de la opsonina con la membrana celular gatilla la formación de vacuolas fagocíticas y activa la enzima NADPH oxidasa la cual comienza a generar radicales libres tales como anión superóxido, ion hidroxilo y peróxido de hidrógeno. Esto sucede cuando se está formando el fagosoma (3). Posteriormente, se produce la fusión del fagosoma con lisosomas celulares constituyéndose el fagolisosoma (4). Al vertir su contenido al fagosoma, estos organelos aportan mieloperoxidasa que genera hipoclorito o hipoyodito a partir del peróxido de hidrógeno. El anión superóxido, el ion hidroxilo, el peróxido de hidrógeno y el hipoclorito son parte de los mecanismos bactericidas oxígeno-dependientes. Actúan sobre la membrana bacteriana produciendo peroxidación de lípidos, rompimiento de proteínas de membrana y de uniones disulfuro entre ellas y formación de uniones cruzadas entre lípidos. Los gránulos específicos y azurófilos de los polimorfonucleares aportan también diversas proteínas con capacidad bacteriostática y bactericida. Entre las primeras se encuentran la lisozima que ataca la mureína de la pared bacteriana y la lactoferrina que priva a las bacterias de un elemento esencial para su vida cual es el fierro. Los gránulos azurófilos aportan las proteínas catiónicas microbicidas CAP 57 y CAP 37 que rompen la membrana externa de bacterias gram-negativas. Además, vierten al fagosoma las enzimas proteolíticas catepsina G y elastasa así como la lisozima. Este grupo de elementos microbicidas conforman los mecanismos bactericidas oxígeno-independientes. Finalmente, las enzimas hidrolíticas de los lisosomas digieren a los microrganismos muertos (5). Durante la fagocitosis se produce frecuentemente liberación de enzimas lisosómicas (6) tales como las proteasas neutras las que contribuyen a la fluidificación de la matriz extracelular. Asimismo aporta mediadores químicos de la inflamación tales como las proteínas catiónicas. Estos mediadores aumentan la permeabilidad vascular en forma directa o induciendo la liberación de histamina en la célula cebada. También son quimiotácticos para monocitos e inhiben el movimiento de otros neutrófilos y eosinófilos. La fagocitosis por macrófagos es similar a la descrita para polimorfonucleares neutrófilos difiriendo de esta en que carecen de mieloperoxidasa. A diferencia de los polimorfonucleares, los macrófagos cumplen una labor fundamental en la respuesta inmune adaptativa al ser células presentadoras de antígeno. |
||||