| Moléculas que participan en el reconocimiento del antígeno | Lámina 15 |
|||
La
fase de inducción de la respuesta inmune se inicia
con el reconocimiento antigénico durante el cual
las células del sistema inmune contactan específicamente
a los epitopos o determinantes antigénicos a través
de los receptores idiotípicos ubicados en su membrana.
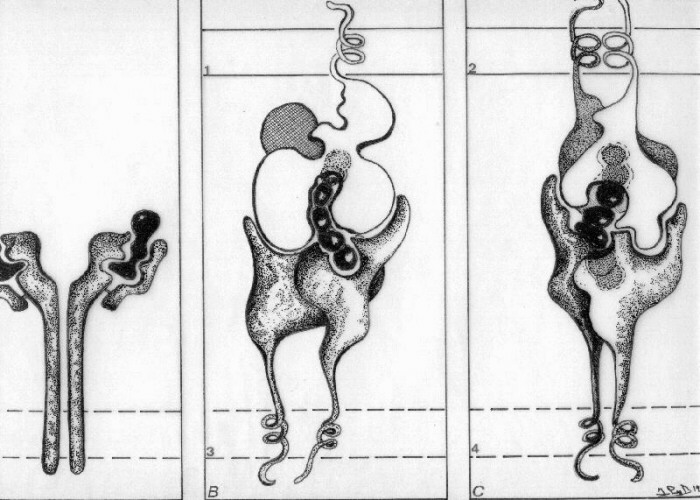
Los linfocitos T reconocen antígeno con restricción genética, esto es, sólo son activados si el péptido antigénico les es presentado por moléculas codificadas por el complejo mayor de histocompatibilidad (MHC). El MHC es una región génica ubicada en el cromosonma 6 humano que codifica para moléculas clase I y clase II y para algunos factores del complemento, citoquinas y ciertas proteínas de shock térmico. En el reconocimiento por linfocitos T CD8+ (b) se requiere de la presencia de moléculas MHC clase I (1). Los linfocitos T CD4+ (c) reconocen antigeno en el contexto de moléculas MHC clase II (2). Los linfocitos reconocen al complejo formado por estas moléculas MHC y el péptido antigénico mediante sus receptores idiotípicos TCR (3 y 4). Los productos MHC clase I son glicoproteínas de membrana
que forman un complejo no covalente con un polipéptido
no polimórfico denominado beta 2 microglobulina que
está codificado en otro gen. Los productos clase
II tienen dos cadenas polimórficas: alfa y beta.
Las estructuras tridimensionales de ambos productos son
similares y pueden ser divididas en cuatro regiones: La zona de los productos clase I y II que une antígeno
presenta una región hipervariable la que a través
de múltiples plegamientos forma una concavidad que
puede acomodar un péptido de 10 a 20 aminoácidos.
Esta región ha sido denominada bolsillo o hendidura. Los receptores TCR presentes en linfocitos T CD8+ y T CD4+ son heterodímeros compuestos por dos cadenas polipeptídicas denominadas alfa y beta. Una pequeña población de linfocitos T presenta un heterodímero formado por cadenas gamma y delta. Estas cadenas tienen gran similitud con las cadenas de inmunoglobulinas al presentar una región variable y una constante. Ambas cadenas están unidas por puentes disulfuro. El TCR tiene una región extracelular que reconoce al péptido antigénico formando complejo con la molécula MHC, una región de transmembrana y una porción citoplasmática. El reconocimiento diferencial de MHC clase I y II por linfocitos T CD8+ y T CD4+ respectivamente no depende del TCR sino de la presencia de moléculas CD8 y CD4 en su membrana. La expresión de moléculas TCR y la función activadora de linfocitos requiere de un conjunto de moléculas adyacentes que forman el complejo CD3 y de moléculas de adhesión celular. |
||||